ユビキチン様タンパク質(ユビキチンようタンパクしつ、英: ubiquitin-like protein、略称: UBL)は、細胞内の他のタンパク質の翻訳後修飾に関与する低分子量タンパク質のファミリーで、多くの場合調節機能を持つ。UBLファミリーの名称は、最初に発見されたメンバーであるユビキチン(Ub)に由来する。ユビキチンは他のタンパク質への共有結合修飾を介してタンパク質分解を調節する役割があることが最もよく知られている。ユビキチンの発見以降、進化的関係のある他のメンバーが多く記載されており、それらもユビキチンと同様の化学反応を伴う並行的な調節過程に関与している。UBLは、オートファジー、タンパク質輸送、炎症および免疫応答、転写、DNA修復、RNAスプライシング、細胞分化など、さまざまな細胞機能に関与している。
発見
ユビキチンは1970年代に発見され、もともとは"ubiquitous immunopoietic polypeptide"と命名されていた。その後、ユビキチンと配列類似性を示す他のタンパク質が報告されたが、タンパク質の共有結合修飾という重要な特徴を共通して持つことが最初に示されたのは、1987年に発見されたISG15である。1990年代半ばになされた一連の報告がこの分野の転換点として認識されており、1996年の同じ時期にさまざまな研究者によってSUMO(small ubiquitin-like modifier、別名: Sentrin、SENP1)が発見され、1997年にはNEDD8、1998年にはATG12が発見された。系統的調査によって、真核生物のゲノムにはユビキチンやユビキチン様タンパク質をコードする遺伝子が10,000種類以上同定されている。
構造と分類
UBLファミリーのメンバーは低分子量の非酵素タンパク質である。約76アミノ酸からなり、ユビキチンに代表されるような、αヘリックスが5本の逆平行βシートで囲まれた"β-grasp"フォールドをとる。β-graspフォールドは、真核生物と原核生物の双方に起源をもつ他のタンパク質にも広く分布している。ユビキチンとユビキチン様タンパク質を総称して「ユビキトン」(ubiquiton)と呼ぶことがある。
UBLは他の分子と共有結合を形成する能力の有無によって、2つのカテゴリに分類される。他の分子と共有結合するUBL(タイプIとも呼ばれる)は、C末端の2つのグリシン残基からなる特徴的な配列モチーフが存在し、このモチーフを介して他の分子への付加が行われる。一般的には、UBLは不活性な前駆体として発現し、C末端のタンパク質分解によって活性グリシンが露出することで活性化される。ほぼすべてのタイプI UBLが最終的には他のタンパク質と連結されるが、少なくとも1つの例外が存在し、ATG8はホスファチジルエタノールアミンに連結される。他の分子と共有結合しないUBL(タイプIIとも呼ばれる)は、より大きなポリペプチド鎖中で他のドメインと融合したタンパク質ドメインとして存在していることが多く、タンパク質分解によってUBLドメインが放出される場合や、タンパク質間相互作用ドメインとして機能する場合がある。より大きなタンパク質に存在するUBLドメインは、UBXドメインとしても知られている。
分布
ユビキチン(ubiquitin)はその名前が示す通り真核生物で普遍的(ubiquitous)に発現しており、細菌や古細菌には存在しないと考えられてきた。しかし、古細菌ではいくつかの例が記載されている。UBLは真核生物で広く分布しているが、その分布は系統によって異なる。例えば、ISG15は免疫系の調節に関与し、下等真核生物には存在しない。他のファミリーでも一部の系統での多様化がみられ、SUMOファミリーのメンバーは酵母のゲノムには1つしか存在しないが、脊椎動物のゲノムには少なくとも4つのメンバーが存在し、その一部には機能的冗長性がみられる。さらに、モデル植物のシロイヌナズナArabidopsis thalianaではゲノム中に少なくとも8つのメンバーが存在する。
ヒト
ヒトゲノムにはユビキチン自身を除いて少なくとも8つのファミリーのUBLがコードされている。他のタンパク質に対する共有結合修飾を行うタイプIと考えられているUBLには、SUMO、NEDD8、ATG8、ATG12、URM1、UFM1、FAT10、ISG15がある。さらに、FUBIと呼ばれるタンパク質はFAU遺伝子に融合タンパク質としてコードされており、タンパク質分解によるプロセシングによって遊離グリシンC末端が生じるが、これが共有結合的タンパク質修飾を行うかどうかは実験的には示されていない。
植物
植物のゲノムはユビキチン以外にSUMO、RUB(NEDD8の植物ホモログ)、ATG8、ATG12、MUB、UFM1、HUB1という7つのUBLのファミリーがコードされていることが知られており、さらに多数のタイプII UBLが存在する。UBLファミリーの一部と関連調節タンパク質は植物で劇的に拡大しており、これは全ゲノム重複と他の形式の遺伝子重複の双方の結果であると考えられている。ユビキチン、SUMO、ATG8、MUBファミリーは植物のUBL遺伝子の約90%を占めると推計されている。ユビキチンやSUMOシグナルの伝達に関係するタンパク質の種類は、有胚植物のゲノムで大きく増加している。
原核生物
真核生物と比較して、原核生物のタンパク質とUBLとの系統学的関係性は限られたものである。放線菌の一部にはPup(prokaryotic ubiquitin-like protein)と呼ばれるタンパク質が存在しており、プロテアソームによる分解のためにタンパク質を標識する点でユビキチンと機能的に類似している。しかしながら、これらは天然変性タンパク質であり、UBLとの進化的関係性は不明である。近年では、一部のグラム陰性菌系統でもUBactと呼ばれる同様のタンパク質が記載されている。対照的に、テルムス属Thermusの細菌にはTtuBと呼ばれる、真核生物のUBLと共通したβ-graspフォールドを持つタンパク質が存在する。TtuBは硫黄のキャリアタンパク質と、タンパク質の共有結合修飾という二重の機能を持つことが報告されている。古細菌では、SAMP(small archaeal modifier protein)と呼ばれるβ-graspフォールドタンパク質が存在し、タンパク質分解においてユビキチンに似た役割を果たしていることが示されている。2011年に同定された未培養古細菌では真核生物のユビキチン経路に対応すると思われる完全な遺伝子セットが同定されており、ユーリアーキオータ、クレンアーキオータ、アイグアーキオータの少なくとも3つの系統には同様のシステムが存在すると考えられている。さらに、一部の病原性細菌は真核生物のUBL経路を模倣するタンパク質を進化させており、宿主細胞のUBLと相互作用してシグナル伝達機能に干渉する。
調節
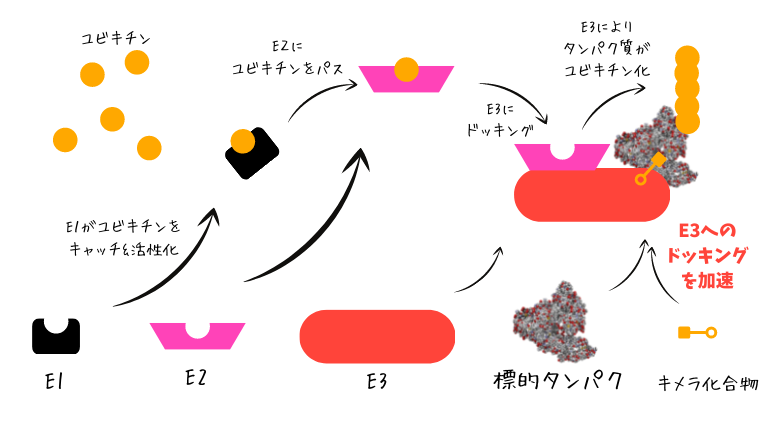
真核生物で共有結合修飾を行うUBLの調節は精巧なものであるが、一般的には各ファミリーごとの平行過程である。こうした調節過程はユビキチンで最もよく特徴づけられている。ユビキチン化の過程は緊密に調節された3つの段階からなる。ユビキチン活性化酵素(E1)による活性化、ユビキチン結合酵素(E2)による結合、ユビキチンリガーゼ(E3)によるライゲーションである。この過程によって、ユビキチンのC末端と標的タンパク質の残基(通常はリジン)の間で共有結合が形成される。多くのUBLファミリーにも同様の3段階過程が存在し、各ファミリーに特異的な酵素のセットによって触媒される。脱ユビキチン化はタンパク質基質からユビキチンを除去する過程であり、脱ユビキチン化酵素(DUB)によって行われる。これらの酵素がUBLに対して作用する範囲はさまざまであり、予測は困難である。SUMOやNEDD8など、一部のUBLにはファミリー特異的なDUBが存在する。
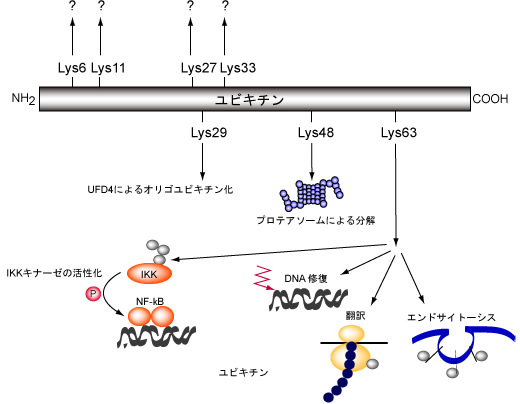
ユビキチンは、タンパク質基質に最初に結合したユビキチン分子にさらにユビキチンが結合することで、多量体の鎖を形成する。こうした鎖は直鎖状である場合も分岐している場合もあり、ユビキチン鎖の長さと分岐によって異なる調節シグナルが送られている可能性がある。鎖の形成はすべてのUBLファミリーで確認されているわけではないが、SUMO、NEDD8、URM1では実験的に検出されている。さらに、ユビキチン自身もUBLによって修飾されることがSUMOとNEDD8で確認されている。異なるUBLファミリー間での交差について最もよく特徴づけられているのは、ユビキチンとSUMOの関係である。
細胞機能
UBLはファミリー全体では非常に広範囲の細胞過程に関与しており、UBLファミリーによってその活性や修飾を行うタンパク質の多様性が異なる。ユビキチンの最もよく知られた機能はプロテアソームによって分解されるタンパク質を特定することであるが、ユビキチン化は、エンドサイトーシスや他の形態のタンパク質輸送、転写や転写因子の調節、細胞シグナル伝達、ヒストン修飾、DNA修復など他の過程にも関与している。他のほとんどのUBLも同じように細胞過程を調節する役割を持つが、その範囲はユビキチンよりもかなり限定されていることが多い。SUMOタンパク質はユビキチンの次に幅広い細胞タンパク質の標的を持ち、転写、DNA修復、細胞ストレス応答などの過程に関与している。NEDD8はCullinタンパク質を調節する役割が最もよく知られており、ユビキチンを介したタンパク質分解の調節を行うが、他の機能も存在する可能性が高い。ATG8とATG12はオートファジーの過程に関与しており、ATG12は既知の基質が2つだけしかない点で、ATG8はタンパク質ではなくリン脂質のホスファチジルエタノールアミンに結合する点でそれぞれ独特である。
進化
UBLがファミリーとして認識されて以降、UBLと調節タンパク質のセットの進化に関心が寄せられている。β-graspフォールドスーパーファミリーの系統学的研究からは、真核生物のUBLは単系統であり、共通の進化的起源を持つことが示唆されている。UBLと相互作用する酵素のカスケードからなるUBL調節システムは、原核生物での補酵素チアミンとモリブドプテリンの生合成経路と共通の進化的起源を持つと考えられている。これらの経路で硫黄転移タンパク質として機能するThiSやMoaDはβ-graspフォールドがUBLと共通しており、これらの経路のThiFやMoeBとユビキチン活性化タンパク質には配列類似性と共通した触媒機構がみられる。興味深いことに、真核生物のタンパク質URM1はUBLと硫黄の双方のキャリアタンパク質として機能し、この進化的関係を裏付ける分子化石として記載されている。
UBLファミリーと関連タンパク質に関する比較ゲノミクス研究からは、UBLシグナル伝達機構は真核生物の最終共通祖先(LECA)では既に発達しており、祖先の古細菌に起源を持つことが示唆されている。この仮説は、一部の古細菌のゲノムにはユビキチン化経路が完全に機能するために必要な遺伝子が存在しているという観察からも支持される。真核生物系統では、動物と植物の双方の系統での多細胞化の起源に対応する、UBLファミリーの2つの異なる多様化イベントが確認されている。
出典

![]()